-
-

Call Us
185-6516-0333
Call Us
185-6516-0333
 Tang_ivf
Tang_ivf

人类卵泡发生需要近1年的时间才能完成始基卵泡生长发育直至排卵(图2-2)的过程。卵泡的发育分为两个时期:非促性腺激素依赖期和促性腺激素依赖期。始基卵泡离开原始卵泡池向初级卵泡转化,经次级卵泡发育至小窦卵泡,需要200天以上的时间。这一阶段的卵泡细胞不表达FSH及LH受体,为非促性腺激素依赖阶段,由卵巢局部的生长因子通过自分泌或旁分泌的形式调控、从次级卵泡经窦卵泡、主导卵泡发育至排卵前卵泡需要经历持续生长期和指数生长期的阶段,共需要85天,跨越了4个月经周期。通常我们所说的卵泡期是指卵泡发育最后的指数生长阶段,一般需时14天,这一时期为促性腺激素依赖阶段,在此阶段卵泡发育受 FSH、LH 和生长因子的调节。
原始卵泡被程序性募集,离开原始卵泡池向初级卵泡转化,开始缓慢生长的过程称为启动募集。部分已启动募集的卵泡对周期性变化的促性腺激素发生应答,开始加速生长,称为周期募集。周期募集的卵泡多达20~30枚,但只有一枚卵泡被选择发育成熟并最终排卵,这取决于卵泡对FSH的敏感性。在黄体末期﹣卵泡早期的过度阶段,随着黄体的衰退,雌孕激素和抑制素A水平下降,对下丘脑及垂体的负反馈减弱,FSH分泌增加,形成一个小波峰,只有处于最高发育阶段的卵泡因具备最高的FSH敏感性,得以选择发育成为优势卵泡(dominant follicle)。优势卵泡的卵泡液积聚,卵泡腔充盈,颗粒细胞和膜细胞迅速增殖并分泌大量雌激素,负反馈作用于垂体和下丘脑,使FSH水平下降,其余进入周期募集的没有被选择的卵泡随之退化闭锁(图2-3)。
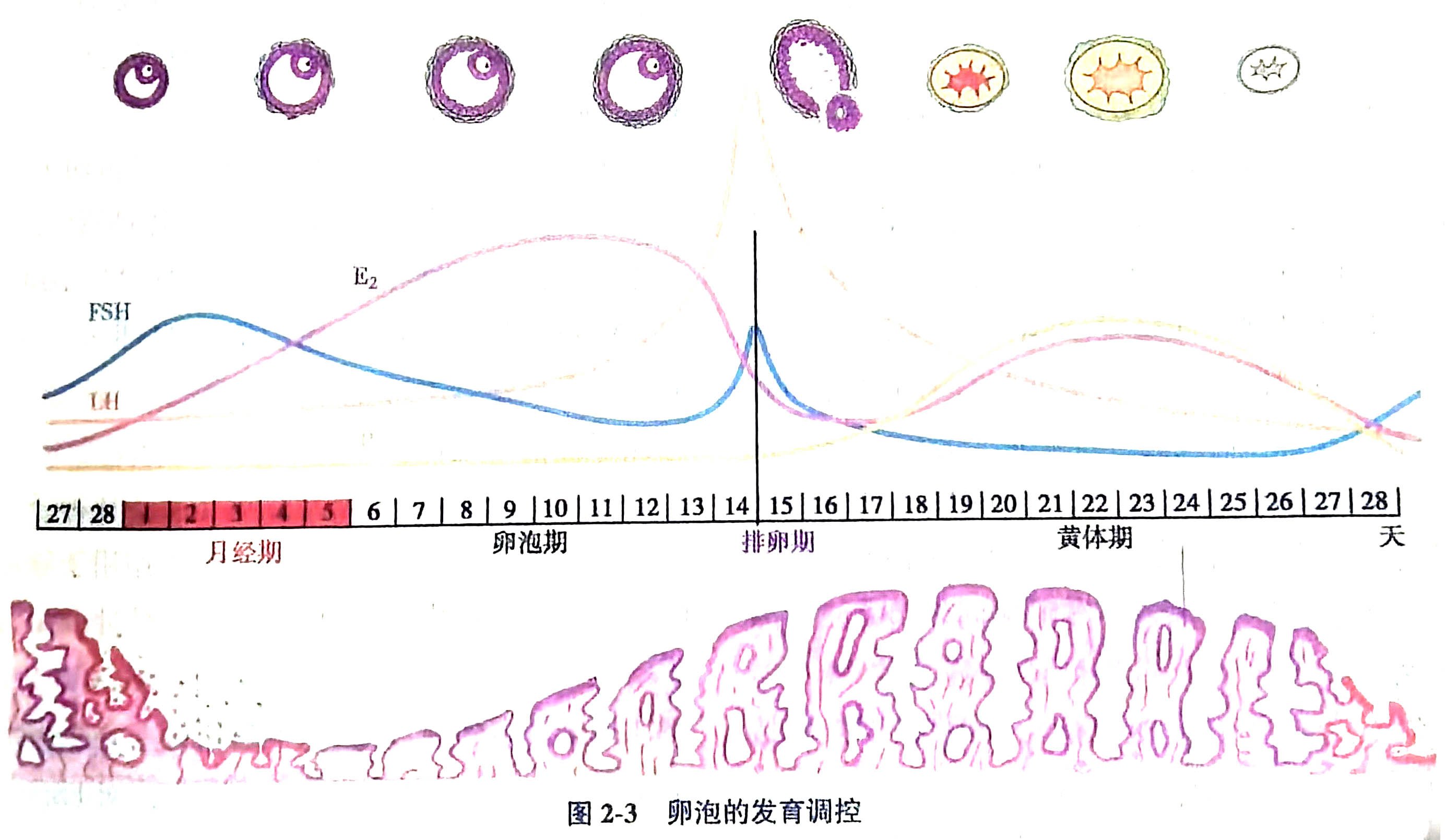
在自然排卵周期中,下丘脑﹣垂体与卵巢(hypothalamus-pituitary-ovary,HPO)互相协调,精密调控,以实现卵泡周期性募集和单卵泡选择。下丘脑向垂体门脉系统脉冲式分泌促性腺激素释放激素(gonadotropin-releasing hormone,GnRH)以调节腺垂体(垂体前叶)合成和释放促性腺激素(gonadotropin, Gn),进而促进卵泡的发育、排卵和黄体形成,调控雌孕激素及抑制素的分泌。而卵巢分泌的甾体激素和抑制素又反馈作用于下丘脑和垂体,调节GnRH和Gn的释放。FSH对中枢的负反馈调控机制保证了自然周期单卵泡发育和排卵,而雌激素诱发的正反馈LH峰是排卵的先决条件。
(一)下丘脑的相关神经内分泌激素
1. GnRH 丘脑位于脑基底部,第三脑室的下方,视交叉和垂体的上方,是生殖内分泌系统的协调中心,能产生多种肽类激素。其中GnRH 是与生殖内分泌关系最为密切的神经肽类激素。
GnRH是8号染色体短臂上基因编码的十肽激素,因结构中的氨基酸连接易被肽酶破坏,因此天然GnRH在血液中的半衰期仅2-4分钟。
合成分泌GnRH的神经元大部分集中在下丘脑内侧基底区、弓状核及下丘脑前部视前区,尤以下丘脑弓状核为主。下丘脑神经元产生的GnRH 经结节漏斗干内的轴突运送至正中隆起,然后释放进入腺垂体(垂体前叶)的垂体门脉系统,通过与腺垂体细胞上的GnRH受体结合,促进合成与释放FSH、LH以维持生殖功能。与FSH相比,GnRH对LH的控制更占据主导地位。FSH除受GnRH调节外,还受激活素、抑制素、卵泡抑制素及抗米勒管激素(AMH)等的调节。
GnRH的释放具有内在的脉冲性和节律性,女性月经周期的不同阶段GnRH脉冲释放频率不同。早卵泡期逐渐升高的雌激素抑制了GnRH分泌脉冲的幅度,对脉冲频率几乎不产生影响,因此早卵泡期呈高频低幅型(90~120分钟1次),而黄体期的高孕激素水平主要抑制GnRH脉冲的频率,因此呈低频高幅型(3-4小时1次)。
2. GnRH分泌的调节 GnRH神经元的分泌受中枢神经系统的调节。同时,多种神经递质、激素和生长因子可交互作用共同调节GnRH的释放。
(1)局部神经递质的调节:儿茶酚胺类(包括多巴胺、去甲肾上腺素、肾上腺素)、B﹣内啡肽、神经肽﹣Y和单胺类物质可通过影响 GnRH 脉冲频率调节 GnRH 的释放。弓状核以及室旁核分泌的多巴胺,可以直接促进下丘脑GnRH神经元的释放,还可以通过垂体门脉系统,抑制垂体分泌PRL,参与HPO轴调控。神经肽﹣Y可以直接作用于GnRH神经元,在高雌激素环境下促进下丘脑GnRH 的脉冲式分泌。
(2)反馈系统的调控:卵巢甾体激素对GnRH 神经元分泌功能的长反馈调节以及腺垂体(垂体前叶)来源的Gn对GnRH神经元分泌功能的短反馈调节已被大家所熟知。近年来,GnRH的超
短反馈调节也受到关注。研究发现,GnRH浓度的变化可反过来作用于下丘脑 GnRH 神经元,通过增减自身受体数来调节自身的分泌。当GnRH 浓度过高或非脉冲式持续刺激时,细胞膜受体内化以减少GnRH受体,降低其生物反应性。GnRH脉冲分泌对于维持稳定的GnRH 受体数目是必需的。
(3)神经调控:中枢神经系统的边缘系统、新皮质、中脑等区域与下丘脑形成复杂的神经网络,体内外的各种刺激均可以通过神经网络干扰下丘脑神经元的脉冲分泌。比如,位于视前交叉区内的 y﹣氨基丁酸神经元(y-aminobutyric acid,GAGB)以及下丘脑KNDy神经元及Kiss1神经元可通过Kisspeptin分泌调节GnRH水平。
(4)AMH 对GnRH的调控: AMH 是由卵巢中窦前、小窦卵泡颗粒细胞分泌的二聚体糖蛋白,其分泌不受促性腺激素及月经周期的影响。AMH 与AMH 受体(AMH receptor,AMHR)-II结合后通过激活果蝇母本抗生存因子蛋白(Smad蛋白)调控下游信号通路。下丘脑表达AMH受体,通过血脑屏障的AMH可直接作用于GnRH神经元的树突及神经轴的终末端,或者通过作用于脑室膜细胞及血管内皮细胞,增加GnRH脉冲分泌的频率,使得LH分泌增多。这也是 PCOS 人群 LH/FSH 比值升高的原因之一。此外,除了在中枢水平发挥作用,AMH还可以降低窦前卵泡对FSH的敏感性,作为始基卵泡生长抑制因子,抑制始基卵泡的启动募集,防止卵泡过快过早消耗,维持卵巢储备。
(二)垂体及其生殖相关激素
垂体位于硬脑膜外,处于视交叉下的蝶鞍内,分为前叶和后叶。在下丘脑 GnRH 脉冲式刺激下,垂体前叶嗜碱性细胞脉冲式地分泌Gn。同时,卵巢激素的反馈调节,以及卵泡细胞的自分泌、旁分泌因子也参与Gn分泌的调控。
1.Gn及Gn受体的结构 Gn包括LH和FSH,两者均为糖蛋白激素,由两个共价键连接的a和B亚基构成。两者a亚基完全相同,有差异的B亚基决定了激素的特异性。
FSH和LH受体均属于G蛋白偶联受体家族,由7个跨膜螺旋组成。FSH、LH与细胞外区域结合会引起受体跨膜区的构象改变,通过Gs﹣腺苷酸环化酶途径,导致细胞内环腺苷酸(cAMP)增加,激活下游信号通路。FSH受体存在于卵巢颗粒细胞,LH受体则存在于卵泡膜细胞和黄体细胞中。在卵泡发育后期,FSH 诱导颗粒细胞同时表达LH受体,称为两细胞两促性腺激素原理。
2.Gn的功能及临床意义
(1)FSH:只有颗粒细胞表达FSH受体,因此FSH只作用于颗粒细胞,通过促进颗粒细胞芳香化酶活性,将膜细胞来源的雄烯二酮转化为雌激素。FSH 还刺激颗粒细胞合成抑制素,一方面抑制垂体FSH的分泌,使非优势卵泡闭锁,实现单卵泡发育,同时还通过旁分泌作用,刺激膜细胞雄激素合成,为晚卵泡期大量雌激素合成提供足够的底物。在周期募集的卵泡波群进一步发育、选择、优势化的过程中,FSH也起到关键诱导作用。在卵泡发育后期,FSH、E2及少量LH诱导卵泡颗粒细胞表达LH受体,从而维持优势卵泡在FSH下降后继续发育直至成熟。LH受体的表达使颗粒细胞能够对LH峰产生反应,促使卵母细胞完成第一次减数分裂,进而卵泡发生排卵。在黄体期,低水平的LH维持卵巢的黄体功能,促进雌孕激素的分泌。
(2)LH:早卵泡期LH只作用于膜细胞,刺激膜细胞17a﹣羟化酶合成雄激素,一方面为颗粒细胞合成雌激素提供底物,另一方面通过旁分泌作用增加颗粒细胞对FSH的敏感性,在早卵泡期发挥促发育的作用,故LH不足可能影响卵泡的成熟。在早卵泡期,适时添加LH,可通过提高雄激素水平,降低FSH阈值,增加募集卵泡数,减少卵泡闭锁的发生。在晚卵泡期,随着颗粒细胞LH受体的形成,LH可同时作用于颗粒细胞和膜细胞,在FSH 下降时代偿部分FSH对颗粒细胞的作用,并为LH峰的到来作好准备。
LH 双相调节17a﹣羟化酶的活性,这种双相调节区别于肾上腺和睾丸,是卵巢所特有的;而LH 对促进胆固醇裂解酶活性的促进作用是单向的,此特点决定了卵泡期的高雌激素和黄体期的高孕激素。卵泡期逐渐上升的LH上调17a﹣羟化酶活性,诱导△5途径有利于雌激素的合成,而在LH峰后,大剂量LH下调17α﹣羟化酶,转向△4途径使雌激素水平下降,孕酮大量合成,故黄体期具有高孕酮水平。
(三)卵巢及其生殖相关激素
卵巢分泌的生殖内分泌类固醇激素主要包括雌激素、孕激素和雄激素,三种激素均以环戊烷并多氢菲的母核为分子骨架。
1.雌激素的功能与临床意义 雌激素是卵巢颗粒细胞分泌的类固醇激素,少部分由肾上腺分泌。天然存在的雌激素按作用强弱依次是雌二醇(estradiol,E2)、雌酮(estrone,E)和雌三醇(estriol,E2)。E3是E,和E2的外周代谢产物。雌激素的主要作用是促进和维持女性生殖器官的发育及第二性征的出现。
(1)在月经周期中,雌激素通过正、负反馈调节HPO轴,参与卵泡发生及排卵的调控。同时,雌激素具有增强FSH效应和诱导颗粒细胞增殖的作用,通过提高卵泡对FSH的敏感性参与优势卵泡的选择,并形成卵泡生长和成熟必要的雌激素微环境;在卵泡早中期,随着卵泡发育和雌激素分泌的增加,FSH受负反馈抑制而下降。只有分泌雌激素能力强的卵泡具有最高FSH敏感性,得以继续发育,被选择为优势卵泡或主导卵泡。
(2)在子宫,雌激素可增加子宫肌层血供,促进子宫平滑肌细胞增生,与子宫肌瘤的发生相关。同时,雌激素促进子宫内膜修复和增生。长期缺乏孕激素拮抗的雌激素刺激可诱发子宫内膜过度增生甚至诱发子宫内膜癌。
(3)促进乳腺导管上皮细胞有丝分裂。青春期后,雌激素水平升高,从而促进乳房发育,女性第二性征表现。
(4)促进女性外阴、阴道、子宫颈和输卵管的发育。雌激素可促进阴道鳞状上皮增生和角化,宫颈腺体分泌增加。
(5)其他:雌激素促进骨骼发育,抑制骨吸收,还参与调节血脂代谢。绝经后妇女雌激素水平下降,导致骨质疏松、心血管疾病风险明显增加。另外,雌激素可以刺激肝脏合成多种凝血因子,如II、VI、IX、X等。雌激素过高,引起凝血功能增强,增加血栓形成风险。
2.孕激素的功能与临床意义 孕激素主要包括孕酮和17a﹣羟孕酮。其中,孕酮主要在黄体卵泡膜细胞和黄体颗粒细胞中合成。排卵后,若次级卵母细胞未受精,则黄体功能衰竭,孕酮分泌减少。孕激素的生理作用包括:
(1)与雌激素类似,可负反馈调节HPO轴,抑制FSH和LH的分泌,参与卵泡发生及排卵的调控。在排卵后,孕激素抑制子宫内膜上皮细胞增殖,使增殖期子宫内膜向分泌期转化,协调子宫内膜与受精卵发育的同步化,为受精卵着床和发育作好准备。
(2)孕激素对子宫平滑肌产生抑制的作用,使子宫平滑肌松弛,张力减弱。在孕期,高水平孕激素一方面通过抑制前列腺素合成,降低肌细胞内钙离子水平,降低子宫对缩宫素的敏感性;另一方面有效抑制母体对胎儿的免疫反应,以维持妊娠。
(3)其他:孕激素能促进乳腺小叶的发育;促进阴道鳞状上皮细胞的脱落,抑制宫颈腺体分泌。另外,孕激素可作用体温中枢,上调体温0.3~0.5℃,这也是基础体温监测可评价排卵的理论基础。
3.雄激素功能与临床意义 卵巢来源的雄激素主要由卵巢膜细胞合成,包括雄烯二酮、少量的睾酮及脱氢表雄酮(dehydroepiandrosterone,DHEA)。在正常女性中,雄烯二酮40%-50%来自卵巢,40%~50%来自肾上腺,其余的来自外周组
织;DHEA主要来源于肾上腺网状带,只有20%的由卵巢膜细胞合成;睾酮则25%来源于肾上腺束状带,25%由卵巢基质直接合成,50%由循环中雄烯二酮在卵巢等肾上腺外组织转化而来。
雄激素是孕烯醇酮合成雌激素过程中的关键中间产物,为雌激素合成提供底物,参与调控多种生理功能:
参与卵泡发育调控:膜细胞产生的雄激素作用于颗粒细胞的雄激素受体,增加颗粒细胞对FSH的敏感性,促进颗粒细胞增殖,形成小窦卵泡前的雄激素环境,支持程序性卵泡募集,阻止卵泡闭锁。异常窦卵泡期后的高雄激素卵巢微环境,一方面直接不利于卵泡发育,导致卵泡闭锁、黄体功能不足;另一方面造成血管生成失调,诱发子宫内膜炎症状态,导致子宫内膜容受性降低。
(2)适宜雄激素水平有助于维持女性的性功能。
(3)参与非生殖功能调节:血管、乳房、皮肤、肌肉、脂肪和骨骼也表达雄激素受体,表明雄激素在外周组织具有作用。雄激素促进外周毛发的生成,蛋白质合成,并参与血脂代谢的调节。
三、排卵的生物学事件
在卵泡发育后期,FSH诱发排卵前颗粒细胞LHR的出现,使得在卵泡成熟的最后阶段,LH的作用较FSH更占优势,促使主导卵泡成熟以准备LH峰的到来。当LH达到某一阈值时,一系列事件发生,包括卵母细胞减数分裂及第一极体的排出、卵泡壁薄化、卵丘颗粒细胞扩张等,最终实现排卵。
(一)第一极体排出
如前所述,自妊娠20周,胚胎卵巢中600万~700万个卵原细胞逐渐进入减数分裂,并停滞在第一次减数分裂双线期,形成初级卵母细胞。初级卵母细胞逐渐被单层梭形前颗粒细胞包围,形成始基卵泡,并进入长达数十年的休眠期。进入青
春期后,下丘脑散发的GnRH脉冲出现,逐渐形成规律的脉冲分泌,HPO轴成熟,雌激素的正反馈诱导垂体LH 峰出现,LH峰的出现导致卵母细胞及卵丘细胞间的缝隙连接关闭,cAMP不能经卵丘细胞进入卵母细胞,卵母细胞局部的cAMP减少,使得卵母细胞局部的卵母细胞成熟抑制因子(oocyte maturation inhibitor,OMI)作用减弱,双线期停滞的初级卵母细胞恢复减数分裂,进而发生卵母细胞核膜溶解,即生发泡破裂(germinal vesicle breakdown,GVBD),这也是减数分裂恢复的第一个形态学标志。
在排卵前,卵母细胞完成第一次减数分裂,产生一个大体积的次级卵母细胞,并排出一个体积小、含另一条同源染色体的细胞,称为第一极体。第一极体的排出是卵母细胞核成熟的标志。第一次减数分裂完成后,次级卵母细胞随即进行第二次减数分裂并停滞在第二次减数分裂中期(MII),直到精子使处于MII期的卵母细胞受精,才能完成第二次减数分裂,并排出第二极体,姐妹染色体单体分离,形成染色体核型为23,X单倍体的卵细胞。这样,一个二倍体生殖细胞经过减数分裂最终产生一个单倍体卵母细胞。
(二)卵泡壁薄化
排卵前LH 峰促使卵泡液剧增,在压力作用下卵泡向卵巢表面突向。随着卵泡体积增大,卵泡壁及卵泡表面卵巢上皮变薄、局部缺血,形成圆形透
明的卵泡小斑。在LH作用下,卵泡液中胶原酶及透明质酸酶表达增加,破坏小斑表面的巢白膜;同时颗粒细胞合成前列腺素增多,一方面促进卵泡壁释放蛋白溶酶,另一方面使卵泡膜外层的卵巢内平滑肌收缩,最终导致小斑破裂。
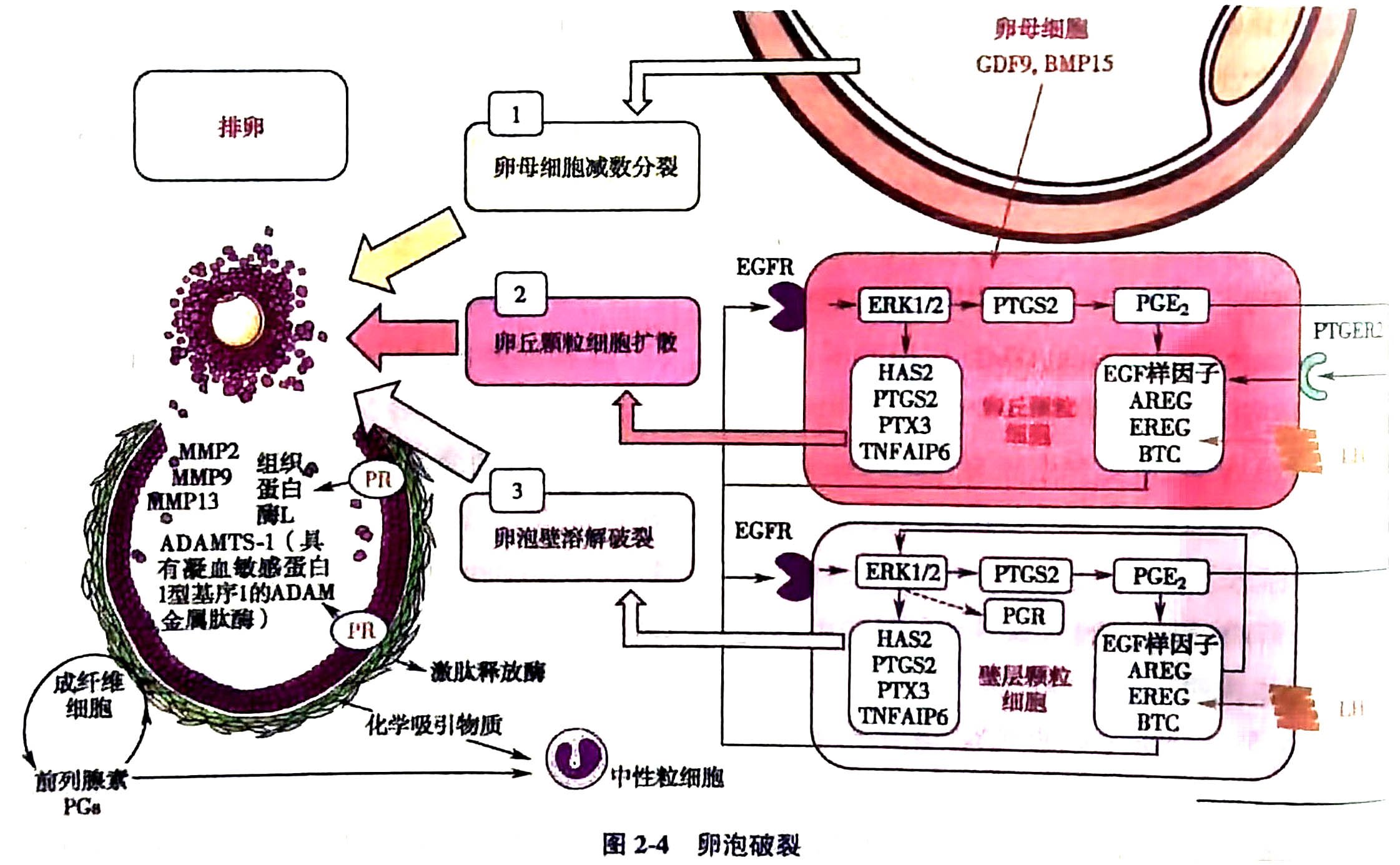
(三)卵丘细胞扩张
排卵前卵泡颗粒细胞分泌高水平雌激素(>200~300pg/ml),持续2~3天,通过kisspeptin神经元介导,正反馈激发LH峰,LH可增加10倍。在LH峰作用下,壁颗粒细胞及卵丘颗粒细胞内的蛋白激酶A (protein kinase A,PKA)激活表皮调节素(epiregulin,EREG),通过与表皮生长因子受体(epidermal growth factor receptor, EGFR)结合激活下游ERK1/2通路,产生前列腺素 E2(prostaglandin E2,PGE2)。PGE2一方面增强平滑肌的收缩,促进COC 排出;另一方面则促进卵丘颗粒细胞表达扩张相关基因,如透明质酸合成酶2(hyaluronan synthase 2,HAS2)、前列腺素内过氧化物合酶2(prostaglandin-endoperoxide synthase 2, PTGS2)、穿透素 3(pentraxin 3,PTX3)、肿瘤坏死因子 a 诱导蛋白6(tumor necrosis factor-alpha induced protein 6,TNFAIP6)的表达,使9卵丘细胞分散,卵丘扩张、黏液化,卵丘复合体与卵泡壁分离并自由悬浮在卵泡液中,最终实现COC 的顺利排出(图2-4)。动物实验中,Bmp-15缺陷雌鼠卵丘膨胀障碍,影响受精而导致不孕,证实卵母细胞源性因子同样参与卵丘扩张的调控。